

Az áramlási viszonyok és a nászruha hatása a test morfológiai jellemzőire
Mozsár Attila1*, Urbán Zsuzsanna2, Pavlikovics Zsuzsanna2, Czeglédi István3, Nyeste Krisztián2, Kati Sára2, Nagy Sándor Alex2, Antal László2
1NAIK Halászati Kutatóintézet, Szarvas
2Debreceni Egyetem, TTK, Hidrobiológiai Tanszék, Debrecen
3MTA Ökológiai Kutatóközpont, Balatoni Limnológiai Intézet, Tihany
*mozsar.attila[kukac]haki.naik.hu
Kivonat
Bevezetés
Számos vizsgálatban olvashatunk az élőhely kínálta környezeti feltételek nyomán kialakuló morfológiai változásokról, illetve a szaporodási időszakra jellemző nászruha morfológiai hatásáról. Arról azonban, hogy e két tényező együttesen, hatásukat egyszerre kifejtve, milyen fajon belüli változásokat okoz a test külső morfológiai jellemzőiben, hiányos ismeretekkel rendelkezünk.
Munkánk során az eltérő környezeti feltételek – áramlási viszonyok – hatására kialakuló, populációk közötti morfológiai variabilitást, és az időszakosan megfigyelhető nászruha okozta populáción belüli különbségeket jellemeztük az amurgéb esetében.
Anyag és módszer
Az amurgéb tág tűrőképességének köszönhetően több élőhelytípusban előfordul, valamint hímjeit feltűnő nászruha (fejdudor) jellemzi a szaporodási időszakban, így kitűnő modell állatként szolgált vizsgálatainkhoz. Mintáinkat egy vízfolyásból (Lónyai-főcsatorna, Tiszabercel) és egy állóvízből (Cserőközi Holt-Tisza, Tiszaderzs) gyűjtöttük havi rendszerességgel, májustól novemberig. A mintavétel során minden esetben az első 50 egyed került begyűjtésre. A halakat szegfűszegolajjal túlaltattuk, és formalinban (5v/v%) tartósítottuk feldolgozásig. A test morfológiai jellemzésére 25 paramétert jelöltünk ki (1. kép).
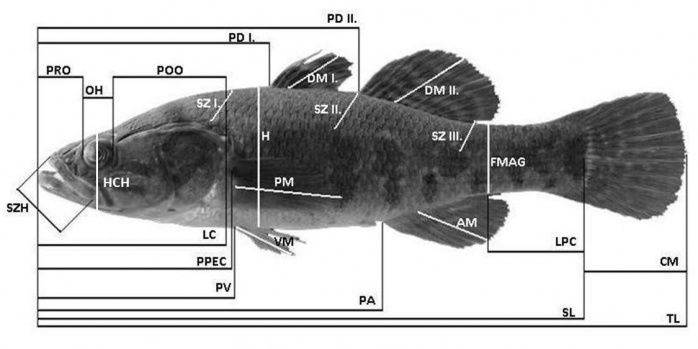
1. ábra: A vizsgálat során mért morfológiai paraméterek
A testméret hatásának kiküszöbölése érdekében az alapadatainkat standardizáltuk (Elliott et al., 1995). Az elemzés első lépéseként morfológiai paraméterenként külön modellekben (három-utas ANOVA) vizsgáltuk az élőhely, az ivar és a mintavételi időpont (mint faktorok) hatását. Ezt követően az összes vizsgált morfológiai paramétert egyszerre figyelembe véve vizsgáltuk az élőhely és az ivar (nászruha) hatását az idő függvényében. Ehhez mintavételi időpontonként diszkriminancia elemzést futtattunk, ahol az a priori csoportokat az élőhelyek és az ivar alapján jelöltük ki.
Eredmények és következtetések
A három-utas ANOVA alapján a testmagasság (H), a második hátúszó hossza (DMII), az anális úszó hossza (AM), a hasúszók hossza (VM), az operculum posterior végénél mért testszélesség (SZ1) és a fejbúb magasság (HCH) értékeire volt lényeges hatással az élőhely, a mintavétel ideje és az ivar. Ezen paraméterek esetében a vizsgálatba vont faktorok a varianciának legalább 30%-át magyarázták (korrigált determinációs együttható, R2 > 0,3).
Az élőhelyek között a testmagasságban és a hasúszó hosszában tapasztaltunk jelentős különbségeket. A vízfolyásban élő populációra, a szakirodalmi adatoknak megfelelően (Webb 1984, McLaughlin és Grant 1994), kisebb testmagasság és nyúlánkabb testforma volt jellemző. A hasúszók esetében az állóvízi populációnál mértünk magasabb értékeket. Ivari különbségeket a második hátúszó és az anális úszó hosszában figyelhettünk meg. A hímeknél ezen úszók hosszabbak, nagyobbak voltak, melyek a testfelületet megnövelve segítik a hímeket a riválisaik, vagy a fészekrablók elijesztésében (Baerends and Baerends van Roon, 1950). A hímek nászruhájaként kialakuló megnövekedett fejbúb a szaporodási időszakban szintén igen jelentős ivarok közötti különbséget eredményezett. A fejbúb egyrészt segít felismerni a hímeket, másrészt információval szolgálhat a hím termékenységéről és szomatikus kondíciójáról (Shaldon 1994, Barlow és Siri 1997). A test szélessége és a magassága az idő függvényében jelentősen változott, feltételezhetően az ivarszervek érésének, és a nyári intenzív tesi növekedés hatására bekövetkező testformaváltozásoknak köszönhetően.
A többváltozós elemzések eredményei alapján a csoportok közötti elkülönülésért elősorban azon morfológiai paraméterek voltak felelősek, melyeknél magas magyarázott varianciát kaptunk az ANOVA modellekben. A tavaszi mintavételeknél jelentős élőhelyek és - a nászruhának köszönhetően - ivarok közötti elválást figyelhettünk meg. Az év előrehaladtával és az időszakosan megjelenő fejdudor visszahúzódásával az ivari különbségek mértéke lecsökkent és a csoportok az élőhelyek között mutattak különbséget és elválást.
A környezeti feltételek hatására kialakuló, populációk közötti különbségek és a nászruha okozta fokozott ivari dimorfizmus egyszerre, egymás mellett megfigyelhető hatásokként jelentkeztek. A populációk és az ivarok közötti különbségekért más-más morfológiai paraméterek voltak felelősek, így a két tényező hatása egymástól elválasztható, párhuzamosan vizsgálható.
Összefoglalás
Munkánk során az eltérő áramlási viszonyok hatására kialakuló, populációk közötti morfológiai variabilitást, és az időszakosan megfigyelhető nászruha okozta populáción belüli különbségeket jellemeztük az amurgéb esetében. Modellállatként az amurgébet választottunk, melynek egy állóvízi és egy folyóvízi populációt mintáztuk májustól novemberig havi rendszerességgel. A halakon 25 morfológiai paramétert mértünk. A testmagasság, a hasúszó hossza az élőhelyek között mutatott különbséget, míg a második hátúszó hossza és a fejbúbmagasság az ivarok között mutatott különbséget. A test szélessége az idő függvényében változott. Eredményeink alapján a nászruhának tulajdonítható fokozott ivari különbségek az élőhely környezeti feltételei hatására kialakuló populációk közötti variabilitás egyszerre, egymást nem elfedve jelenek meg.
Kulcsszavak: morfometria, amurgéb, Perccottus glenii, fejdudor
Köszönetnyilvánítás
Köszönetünket szeretnénk kifejezni a mintavételben segédkezőknek és a DE TTK Hidrobiológiai Tanszéknek és munkatársainak, hogy a munkánkhoz szükséges feltételeket biztosították.
Irodalom
Baerends G.P.; Baerends-Van Roon J.M. 1950. An introduction to the study of the ethology of the cichlid fishes. Behaviour. Supplement, III-243.
Barlow G.W.; Siri P. 1997. Does sexual selection account for the conspicuous head dimorphism in the Midas cichlid? Anim. Behav. 53(3), 573-584.
Elliott NG, Haskard K, Koslow JA. 1995. Morphometric analysis of orange roughy (Hoplostethus atlanticus) off the continental slope of southern Australia. J. Fish. Biol. 46, 202–220.
McLaughlin R.L.; Grant J.W.A 1994. Morphological and behavioral differences among recently-emerged brook charr, Salvelinus fontinalis, foraging in slow- vs. fast-running water. Environmental Biology of Fishes, 39(3), 289–300.
Webb P.W. 1984. Body form, locomotion and foraging in aquatic vertebrates. American Zoologist, 24(1), 107-120.
Programajánló
Jelenleg nincs aktuális esemény.
Hírek
2021. január 29., péntek, 16:22
Tisztelt Látogatók!
A hazai agrár-felsőoktatás szükséges megújulásának mérföldköve az alapítványi fenntartású Magyar Agrár- és Élettudományi Egyetem (MATE) létrejötte, amely 2021. február 1-től 5 campuson, több mint 13 ezer hallgató számára fogja össze a dunántúli és közép-magyarországi élettudományi és kapcsolódó képzéseket. Az intézményhez csatlakozik a Nemzeti Agrárkutatási és Innovációs Központ (NAIK) 11 kutatóintézete is, így az új intézmény nem csupán egy oktatási intézmény lesz, hanem az ágazat szellemi, szakpolitikai és innovációs központjává válik, amely nagyobb mozgásteret biztosít a képzések, a gazdálkodás és szervezet modernizálásához, fejlesztéséhez. Az összeolvadással magasabb fokozatra kapcsolunk, a kutatói és egyetemi szféra szorosabban fonódik majd össze, aminek következtében még több érdekes, izgalmas kutatás-fejlesztés születhet majd az agrárium területén.
Kérjük, kövesse tevékenységünket a jövőben is a www.uni-mate.hu honlapon!

2020. október 6., kedd, 08:34
A szokásostól eltérően az idei évben ősszel, október 03-04 között került megrendezésre az Ultrabalaton csapatversenye. NAIK-os csapat az idei évben állt először rajthoz a 14. alkalommal kiírt versenyen.